Lystrosaurus sp.
Ill.: Dmitry Bogdanov
(Da; www.wikipedia.it)
Phylum: Chordata Haeckel, 1874
Subphylum: Vertebrata Cuvier, 1812
Classe: Synapsida Osborn, 1903
Ordine: Therapsida Broom, 1905
Famiglia: Lystrosauridae Broom, 1903
Genere: Lystrosaurus Cope, 1870
Specie e sottospecie
Il genere annovera attualmente le seguenti specie: Lystrosaurus curvatus Owen, 1876 - Lystrosaurus declivis Owen, 1860 - Lystrosaurus maccaigi Seeley, 1898 - Lystrosaurus murrayi Huxley, 1859 - Lystrosaurus declivis Owen, 1860.
Come spesso accade con i fossili, c'è parecchio dibattito nella comunità paleontologica su quante specie siano effettivamente presenti nel bacino del Karoo. Studi condotti dagli anni '30 agli anni '70 suggerivano un numero elevato, persino 23 in uno dei casi.[8] Tuttavia, negli anni '80 e '90, dopo un'attenta analisi degli esemplari a loro disposizione, il numero di specie valide proveniente dal Karoo è stato ridotto a 6: Lystrosaurus curvatus, Lystrosaurus platyceps, Lystrosaurus oviceps, Lystrosaurus maccaigi, Lystrosaurus murrayi e Lystrosaurus declivis. Uno studio del 2011 ha ridotto quel numero a quattro, trattando i fossili precedentemente etichettati come il Lystrosaurus platyceps e il Lystrosaurus oviceps come membri della specie Lystrosaurus curvatus. Il Lystrosaurus maccaigi è la specie più grande e apparentemente più specializzata, mentre il Lystrosaurus curvatus era la meno specializzata. Un fossile simile al Lystrosaurus, il Kwazulusaurus shakai, è stato anch'esso ritrovato in Sudafrica. Sebbene non sia stato assegnato allo stesso genere, il Kwazulusaurus shakai è molto simile al Lystrosaurus curvatus. Alcuni paleontologi hanno quindi proposto che il Kwazulusaurus shakai fosse probabilmente un antenato o un taxon strettamente imparentato con gli antenati del Lystrosaurus curvatus, mentre la specie Lystrosaurus maccaigi derivava da una diversa linea evolutiva. Il Lystrosaurus maccaigi si trova solo nei sedimenti del periodo Permiano e apparentemente non sopravvisse all'evento di estinzione Permiano-Triassico. Le sue caratteristiche specializzate e la sua improvvisa comparsa nel registro fossile senza un antenato ovvio potrebbero indicare che fosse emigrato nel Karoo da un'area in cui non sono stati ritrovati sedimenti risalenti al Permiano superiore. Il Lystrosaurus curvatus si trova in una fascia relativamente stretta di sedimenti risalenti a poco prima e dopo l'estinzione e può essere utilizzato come marcatore approssimativo per il confine tra i periodi Permiano e Triassico. Un cranio identificato come Lystrosaurus curvatus è stato ritrovato nei sedimenti del tardo Permiano dello Zambia. Per diversi anni si è pensato che non ci fossero esemplari Permiani del Lystrosaurus curvatus nel Karoo, il che ha portato a suggerire che il Lystrosaurus curvatus fosse immigrato dallo Zambia nel Karoo. Tuttavia, un riesame degli esemplari Permiani nel Karoo ne ha identificati alcuni come il Lystrosaurus curvatus e non c'è bisogno di supporre un'immigrazione della specie. Le specie Lystrosaurus murrayi e Lystrosaurus declivis sono state ritrovate solo in sedimenti del Triassico. I fossili di Lystrosaurus georgi sono stati ritrovati nei sedimenti del Triassico inferiore del bacino di Mosca, Russia. Era probabilmente strettamente imparentato con il Lystrosaurus curvatus africano, che è considerato una delle specie meno specializzate ed è stato ritrovato nei sedimenti del Permiano superiore e del Triassico inferiore. La specie Lystrosaurus murrayi, oltre a due specie non descritte attualmente assegnate a Lystrosaurus curvatus e Lystrosaurus declivis, è noto anche dalla Formazione Panchet del Triassico inferiore della valle di Damodar e dalla Formazione Kamthi del bacino di Pranhita-Godavari in India. Sette specie di Lystrosaurus sono state descritte dalle formazioni Jiucaiyuan, Guodikeng e Wutonggou del Triassico inferiore dei Monti Bogda nello Xinjiang, Cina, sebbene sia possibile che solo due (Lystrosaurus youngi e Lystrosaurus hedini) siano valide; insolitamente, non sono noti esemplari cinesi di Lystrosaurus al di sotto del confine Permiano-Triassico in questa regione. Le specie Lystrosaurus curvatus, Lystrosaurus murrayi e Lystrosaurus maccaigi sono note dalla Formazione Fremouw nei Monti Transantartici dell'Antartide.
Descrizione
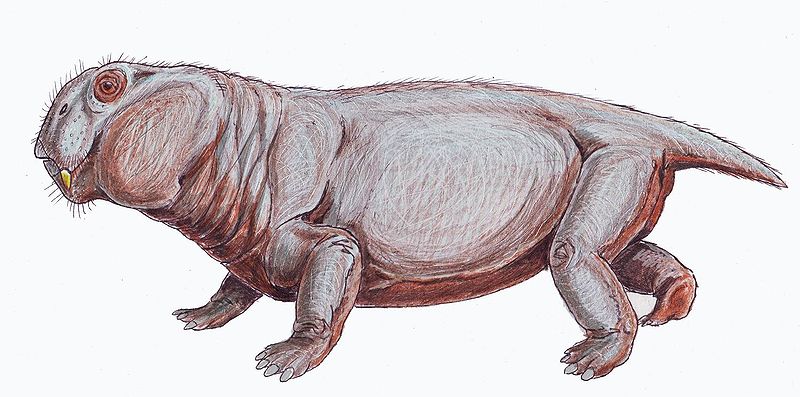
Il dottor Elias Root Beadle, un missionario di Filadelfia e appassionato collezionista di fossili, scoprì il primo cranio di Lystrosaurus. Beadle scrisse della scoperta all'eminente paleontologo Othniel Charles Marsh, ma non ricevette risposta. Il rivale di Marsh, Edward Drinker Cope, si rivelò invece molto interessato alla scoperta e descrisse e nominò l'esemplare fossile come Lystrosaurus nel Proceedings of the American Philosophical Society, nel 1870. Il nome del genere Lystrosaurus deriva dalle parole greche antiche listron ossia "pala" e sauros ossia "lucertola", in riferimento alla forma del cranio vagamente simile alla forma di una pala. In seguito Marsh acquistò il cranio nel maggio 1871, sebbene il suo interesse per un esemplare già descritto non fosse chiaro; ciò potrebbe essere dovuto al voler esaminare attentamente la descrizione e l'illustrazione del suo rivale. La scoperta di fossili di Lystrosaurus a Coalsack Bluff nei Monti Transantartici da parte di Edwin H. Colbert e del suo team nel 1969-1970 ha contribuito a sostenere l'ipotesi della tettonica a placche e a rafforzare la teoria, poiché fossili di Lystrosaurus erano già stati ritrovati in depositi risalenti al Triassico inferiore dell'Africa meridionale, così come in India e in Cina. La maggior parte dei fossili di Lystrosaurus sono stati trovati nelle formazioni Balfour e Katberg del bacino del Karoo, Sudafrica; questi esemplari offrono le migliori prospettive sull'identificazione delle specie perché sono le più numerosi e sono state studiate per più tempo. Il Lystrosaurus era un terapside dicynodonte erbivoro di piccole-medie dimensioni, con una lunghezza compresa tra i 0,6 e 2,5 m, con una media di circa 0,9 m a seconda della specie. A differenza di altri terapsidi, i dicynodonti avevano musi molto corti e schiacciati ed erano privi di dentatura, fatta eccezione per i canini superiori simili a zanne. Il loro corto muso terminava in un becco corneo, simile a quello delle tartarughe, che puntava verso il basso, adatto per tagliare e fare a pezzi la vegetazione, che veniva poi macinata contro un palato secondario corneo quando la bocca si chiudeva. L'articolazione della mascella era debole e si muoveva solamente avanti e indietro con un'azione di taglio, invece dei più comuni movimenti laterali o su e giù presenti nei mammiferi odierni. Si pensa che i muscoli della mascella fossero attaccati insolitamente in avanti sul cranio e occupassero molto spazio sulla parte superiore e posteriore del cranio. Di conseguenza, gli occhi erano posizionati in alto e in avanti sul cranio. Le caratteristiche dello scheletro indicano che il Lystrosaurus si muoveva con un'andatura semi-tentacolare. L'angolo posteriore inferiore della scapola (lama scapolare) era fortemente ossificato, il che suggerisce che il movimento della scapola contribuisse alla lunghezza del passo degli arti anteriori e riduceva la flessione laterale del corpo. Le cinque vertebre sacrali erano massicce ma non fuse tra loro e con il bacino, rendendo la schiena dell'animale più rigida e riducendo la flessione laterale mentre quest'ultimo camminava. Si pensa che i terapsidi con meno di cinque vertebre sacrali avessero arti tentacolari, come quelli delle lucertole moderne. Nei dinosauri e nei mammiferi, che hanno arti eretti, le vertebre sacrali sono fuse tra loro e con il bacino.[16] Si pensa che un contrafforte sopra ogni acetabolo (cavità dell'anca) impedisse la lussazione del femore (osso della coscia) mentre il Lystrosaurus procedeva con un'andatura semi-tentacolare. Gli arti anteriori di Lystrosaurus erano massicci e si pensa che uniti alle grandi zanne al muso piatto dell'animale, queste rendessero il Lystrosaurus un eccellente scavatore. Esemplari mummificati recuperati dal bacino del Karoo e rivelati nel 2022 hanno rivelato che il Lystrosaurus aveva una pelle con senza squame, coriacea e priva di peli. Il genere Lystrosaurus è noto per aver dominato la Pangea meridionale per milioni di anni durante il Triassico inferiore. Almeno una specie non identificata di questo genere sopravvisse all'estinzione di massa della fine del Permiano e, in assenza di predatori e concorrenti erbivori, continuò a prosperare e a irradiarsi nuovamente in un certo numero di specie all'interno del genere,] divenendo il gruppo più comune di vertebrati terrestri durante il Triassico inferiore; per milioni di anni, il 95% dei vertebrati terrestri era composto dal Lystrosaurus. Questa è l'unica istanza in cui una singola specie o genere di animale terrestre ha dominato la Terra a tal punto. Altri pochi generi di terapsidi del Permiano sopravvissero all'estinzione di massa e compaiono nelle rocce del Triassico, come i terocefali Tetracynodon, Moschorhinus, Ictidosuchoides e Promoschorhynchus, ma nessuno di loro raggiunse lo stesso livello di abbondanza durante il Triassico; Il completo recupero ecologico del pianeta durò 30 milioni di anni, abbracciando il Triassico inferiore e medio, fino alla comparsa dei primi dinosauri. Sono state proposte diverse teorie ed ipotesi per spiegare come il Lystrosaurus possa essere sopravvissuto all'evento di estinzione del Permiano-Triassico, la "madre di tutte le estinzioni di massa", ed il come arrivò a dominare la fauna del Triassico inferiore in misura così senza precedenti. I segni di crescita sulle zanne fossilizzate suggeriscono che i Lystrosaurus vissuti in Antartide, circa 250 milioni di anni fa, erano in grado di entrare in uno stato prolungato di torpore analogo al letargo. Questa potrebbe essere la prova più antica di uno stato simile al letargo in un animale vertebrato e indica che il torpore è sorto nei vertebrati prima che mammiferi e dinosauri si evolvessero. Una delle teorie più recenti è che l'evento di estinzione abbia ridotto il contenuto di ossigeno nell'atmosfera e aumentato il contenuto di anidride carbonica, rendendo l'aria più difficoltosa da respirare. È stato quindi suggerito che il Lystrosaurus sia sopravvissuto e abbia raggiunto il suo stato di erbivoro dominante poiché il suo stile di vita da scavatore lo rese in grado di far fronte a un'atmosfera di "aria stantia" e che caratteristiche specifiche della sua anatomia facessero parte di questo adattamento: un torace a botte che ospitava grandi polmoni, narici interne corte che facilitavano una respirazione rapida e alte spine neurali (proiezioni sul lato dorsale delle vertebre) che davano maggiore leva ai muscoli per espandere e contrarre il torace. Tuttavia, ci sono delle lacune in questa teoria: il torace del Lystrosaurus non era significativamente più grande in proporzione alle sue dimensioni rispetto ad altri dicynodonti che si estinsero durante questo periodo; sebbene i dicynodonti del Triassico sembrino aver avuto spine neurali più lunghe rispetto alle loro controparti del Permiano, questa caratteristica potrebbe essere correlata alla postura, alla locomozione o persino alle dimensioni del corpo piuttosto che all'efficienza respiratoria; le specie Lystrosaurus murrayi e Lystrosaurus declivis erano molto più abbondanti di altri scavatori del Triassico inferiore, come il Procolophon o il Thrinaxodon. L'ipotesi che il Lystrosaurus sia sopravvissuto grazie ad uno stile di vita semi-acquatico è stata anch'essa criticata per varie ragioni: l'idea che fosse un animale semi-acquatico è un'ipotesi vecchia dimenticata a favore dell'idea che questi fossero animali fossori o semi-fossori; sebbene i temnospondyli acquatici divennero più abbondanti nei sedimenti triassici del Karoo, erano molto meno numerosi dei Lystrosaurus murrayi e Lystrosaurus declivis. Come regola gli animali più grandi e specializzati sono quelli a più alto rischio durante le estinzioni di massa; questo potrebbe spiegare perché la specie poco specializzata Lystrosaurus curvatus sopravvissero mentre la ben più grande e specializzata Lystrosaurus maccaigi perì insieme a tutti gli altri grandi erbivori e carnivori del Permiano. Sebbene il Lystrosaurus sembri generalmente adattato a nutrirsi di piante simili al Dicroidium, che dominava il Triassico inferiore, le dimensioni maggiori del Lystrosaurus maccaigi potrebbero averlo costretto a fare affidamento sui membri più grandi della flora Glossopteris, che non sopravvissero all'estinzione di fine Permiano. A seguito dell'estinzione del Permiano, molti dei carnivori di grossa taglia presenti nel Permiano si estinsero. Solo il terocefale Moschorhinus, lungo 1,5 metri, e il grande arcosauriforme il Proterosuchus sembrano essere abbastanza grandi da essere in grado di predare le specie di Lystrosaurus triassiche, e questa carenza di predatori potrebbe essere stata responsabile del boom della popolazione del Lystrosaurus nel Triassico inferiore. Secondo Benton, "Forse la sopravvivenza del Lystrosaurus è stata semplicemente una questione di fortuna". Il Lystrosaurus come tutti i sinapsidi preistorici vissuti prima dei dinosauri (escluso il Dimetrodon) è stato poco noto, quasi sconosciuto nella cultura popolare, il suo successo è dovuto in parte grazie al videogioco Ark: Survival Evolved (2017), dove può essere addomesticato, e alla sua apparizione in Jurassic World - Il dominio (2022), film in cui compare un esemplare (soprannominato da alcuni fan Leonard) che non ottenne un ruolo centrale, ma compare per alcuni secondi al centro dellattenzione degli spettatori (soprattutto in un segmento presente nell'edizione estesa del film, dove decapita con le sue fauci un Oviraptor durante un combattimento clandestino nel mercato nero maltese). Prima di allora il Lystrosaurus era apparso nei documentari L'impero dei mostri - La vita prima dei dinosauri (2005), Earth: Making of a Planet (2011) e NHK Leaps in Evolution (2015). Successivamente è apparso sulla serie documentario di Netflix La vita sul nostro pianeta (2023).
Diffusione
I fossili di Lystrosaurus sono stati ritrovati in molti giacimenti ossei terrestri risalenti al Permiano superiore e all'inizio del Triassico, più abbondantemente in Africa e, in misura minore, in parti di quelle che oggi sono India, Cina, Mongolia, Russia europea e Antartide (che all'epoca non si trovava molto più vicino all'equatore).
Bibliografia
(EN) Michael Albert Cluver, Fossil reptiles of the South African Karoo, The South African Museum, 1978.
R.J. Damiani, J. Neveling, S.P. Modesto e A.M. Yates, Barendskraal, a diverse amniote locality from the Lystrosaurus assemblage zone, Early Triassic of South Africa, in Palaeontologia Africana, vol. 39, 2004, pp. 53-62.
Wallace, David Rains, The Bonehunters' Revenge: Dinosaurs, Greed, and the Greatest Scientific Feud of the Gilded Age, Houghton Mifflin Harcourt, 2000, pp. 44-45.
Henry George Liddell e Robert Scott (a cura di), A Greek-English Lexicon, abridged, Oxford, UK, Oxford University Press, 1980.
Naomi Lubick, Investigating the Antarctic, Geotimes, American Geosciences Institute, febbraio 2005. Ospitato su agiweb.org.
(EN) Steve Trewick, Plate Tectonics in Biogeography, in International Encyclopedia of Geography: People, the Earth, Environment and Technology, John Wiley & Sons, Ltd, 2016, pp. 1-9.
M.V. Surkov, N.N. Of Kalandadze e M.J. Benton, Lystrosaurus georgi, a dicynodont from the Lower Triassic of Russia (PDF), in Journal of Vertebrate Paleontology, vol. 25, n. 2, giugno 2005, pp. 402-413.
J. Botha e R.M.H. Smith, Lystrosaurus species composition across the PermoTriassic boundary in the Karoo Basin of South Africa, in Lethaia, vol. 40, n. 2, 2005, pp. 125-137.
F.E. Grine, C.A. Forster, M.A. Cluver e J.A. Georgi, Amniote Paleobiology: Perspectives on the evolution of mammals, birds, and reptiles, University of Chicago Press, 2006, pp. 432-503.
(EN) Saswati Bandyopadhyay e Sanghamitra Ray, Gondwana vertebrate faunas of India: Their diversity and intercontinental relationships, in Episodes Journal of International Geoscience, vol. 43, n. 1, 1º marzo 2020, pp. 438-460.
Kenneth D. Angielczyk, Jun Liu, Christian A. Sidor e Wan Yang, The stratigraphic and geographic occurrences of Permo-Triassic tetrapods in the Bogda Mountains, NW China Implications of a new cyclostratigraphic framework and Bayesian age model, in Journal of African Earth Sciences, vol. 195, 1º novembre 2022, pp. 104655.
(EN) Fenglu Han, Qi Zhao e Jun Liu, Preliminary bone histological analysis of Lystrosaurus (Therapsida: Dicynodontia) from the Lower Triassic of North China, and its implication for lifestyle and environments after the end-Permian extinction, in PLoS One, vol. 16, n. 3, 18 marzo 2021, pp. e0248681.
J.W. Cosgriff, W.R. Hammer e W.J. Ryan, The Pangaean reptile, Lystrosaurus maccaigi, in the Lower Triassic of Antarctica, in Journal of Paleontology, vol. 56, n. 2, 1982, pp. 371-385.
(EN) Fossils & Facts, in Lystrosaurus, Encyclopaedia Britannica.
R. Cowen, The History of Life, 3rd, Blackwell Scientific, 2000, pp. 167-168.
Michael J. Benton, Origin and relationships of Dinosauria, in David B. Weishampel, Peter Dodson e Halszka Osmólska (a cura di), The Dinosauria, 2nd, Berkeley, CA, University of California Press, 2004, pp. 7-19.
Smith, R.M.H., Botha, J. e Vigilietti, P.A., Taphonomy of drought afflicted tetrapods in the Early Triassic Karoo Basin, South Africa, in Palaeogeography, Palaeoclimatology, Palaeoecology, vol. 604, 2022.
These fossil mummies reveal a brutal world long before T. rex lived, su gizmodo.com, 20 settembre 2022.
M.J. Benton, When Life Nearly Died: The greatest mass extinction of all time, Londra, UK, Thames & Hudson, 2006.
Christopher Cokinos, The consolations of extinction, in Orion, 12 ottobre 2007.
A.K. Huttenlocker, Sidor, C.A. e Smith, R.M.H., A new specimen of Promoschorhynchus (Therapsida: Therocephalia: Akidnognathidae) from the Lower Triassic of South Africa and its implications for theriodont survivorship across the Permo-Triassic boundary, in Journal of Vertebrate Paleontology, vol. 31, n. 2, 2011, pp. 405-421.
S. Sahney e M.J. Benton, Recovery from the most profound mass extinction of all time, in Proceedings of the Royal Society B: Biological Sciences, vol. 275, n. 1636, 2008, pp. 759-65.
D.H. Erwin, The Great Paleozoic Crisis: Life and death in the Permian, Columbia University Press, 1993.
(EN) Fossil evidence of 'hibernation-like' state in 250 million-year-old Antarctic animal, su phys.org, agosto 2020.
(EN) Fossil suggests animals have been hibernating for 250 million years, in UPI.
(EN) Megan R. Whitney e Christian A. Sidor, Evidence of torpor in the tusks of Lystrosaurus from the Early Triassic of Antarctica, in Communications Biology, vol. 3, n. 1, 27 agosto 2020, pp. 471.
Surkov, M.V., Kalandadze, N.N., and Benton, M.J. (June 2005). "Lystrosaurus georgi, a dicynodont from the Lower Triassic of Russia" (PDF). Journal of Vertebrate Paleontology 25 (2): 402-413.
Grine, F.E., Forster, C.A., Cluver, M.A. & Georgi, J.A. (2006), "Cranial variability, ontogeny and taxonomy of Lystrosaurus from the Karoo Basin of South Africa", Amniote paleobiology. Perspectives on the Evolution of Mammals, Birds, and Reptiles,, University of Chicago Press, pp. 432-503.
Botha, J., and Smith, R.M.H. (2007). "Lystrosaurus species composition across the PermoTriassic boundary in the Karoo Basin of South Africa". Lethaia 40: 125-137.

|
Data: 24/03/1999
Emissione: Animali preistorici Stato: Central African Republic Nota: Emesso in un foglietto di 6 v. diversi |
|---|